If there is a traditional disciplinary 'glue' at the core of my research, it would probably be freshwater biogeography and macroecology. My interest in these topics arose from my consulting days, when I was intrigued by the pronounced biological differences that I observed in rivers and streams that were geographically far removed, yet characterized by similar physical habitats. As a fisheries scientist, I assumed that extant communities were a direct reflection of their contemporary environments. And so the largely historical world of biogeography became a real eye-opener for me.
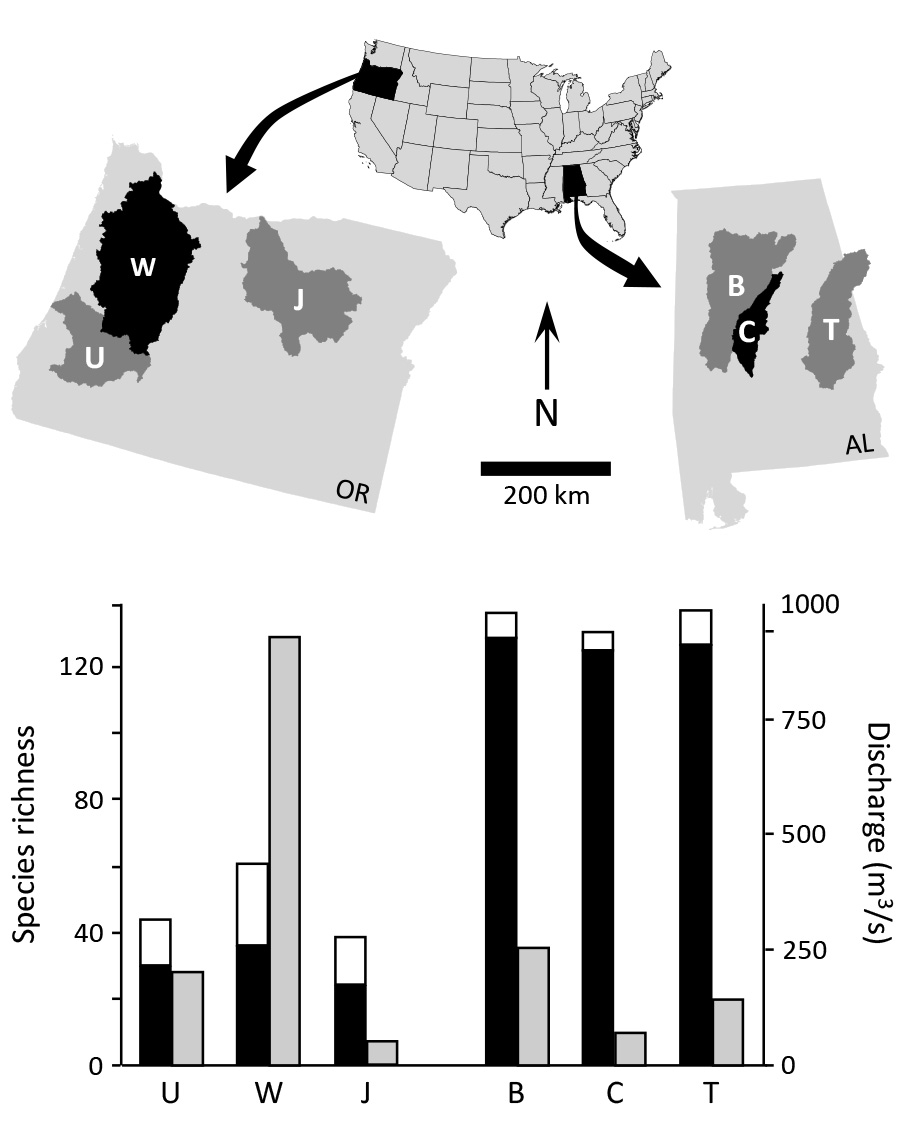
Consider, for example, the disparity in fish species richness between Southeast and Pacific Northwest rivers, as illustrated below. As a consultant, I knew that river size is a strong correlate of fish richness and I was puzzled by the low diversity of the (large) Pacific Northwest rivers that I was working in, relative to the highly diverse Southeast rivers.
Three Alabama rivers ('B' - Black Warrior; 'C' - Cahaba; 'T' - Tallapoosa) and three Oregon rivers ('U' - Umpqua; 'W' - Willamette; 'J' - John Day) are shown. In general, the Pacific Northwest rivers are as large or larger than the Southeast rivers, as indicated by mean annual discharge (gray bars in the lower graph). However, the numbers of native fish species (black bars) are much higher in Southeast rivers. Note also that more non-native fishes (white bars) are established in the Northwest rivers.
One of the first biogeographical / macroecological research objectives that I set my sights on was an improved, operational definition of the 'regional species pool' (RSP) for river fishes. RSP's are fundamental units in biogeography, macroecology and community ecology, and are defined as "groups of potentially interacting species that are physically capable of colonizing and surviving within a given area or habitat." This is important because the absence of a given species within a suitable, potential habitat may be a sign of inter-specific exclusion.
As I came up to speed on my biogeographical reading, it occurred to me that 'ichthyofaunal river zones' may be an ideal means of delineating RSP's within lotic ecosystems. When examined at large spatial scales, such as complete river basins, many riverine fish assemblages can be divided into a series of adjacent, zonal sub-assemblages with high species turnover at zone boundaries and generally homogenous composition within zones. Thus, ichthyofaunal zones consist of co-occurring species within relatively uniform environments: a good functional fit to the conceptual RSP definition.
I then set about using large, composite fish data sets to quantify zonal structure within North American rivers. So far, I've done this for rivers in the Southeast, Pacific Northwest, and the Southwest. An example is shown below for the Rio Chama and Rio Grande.
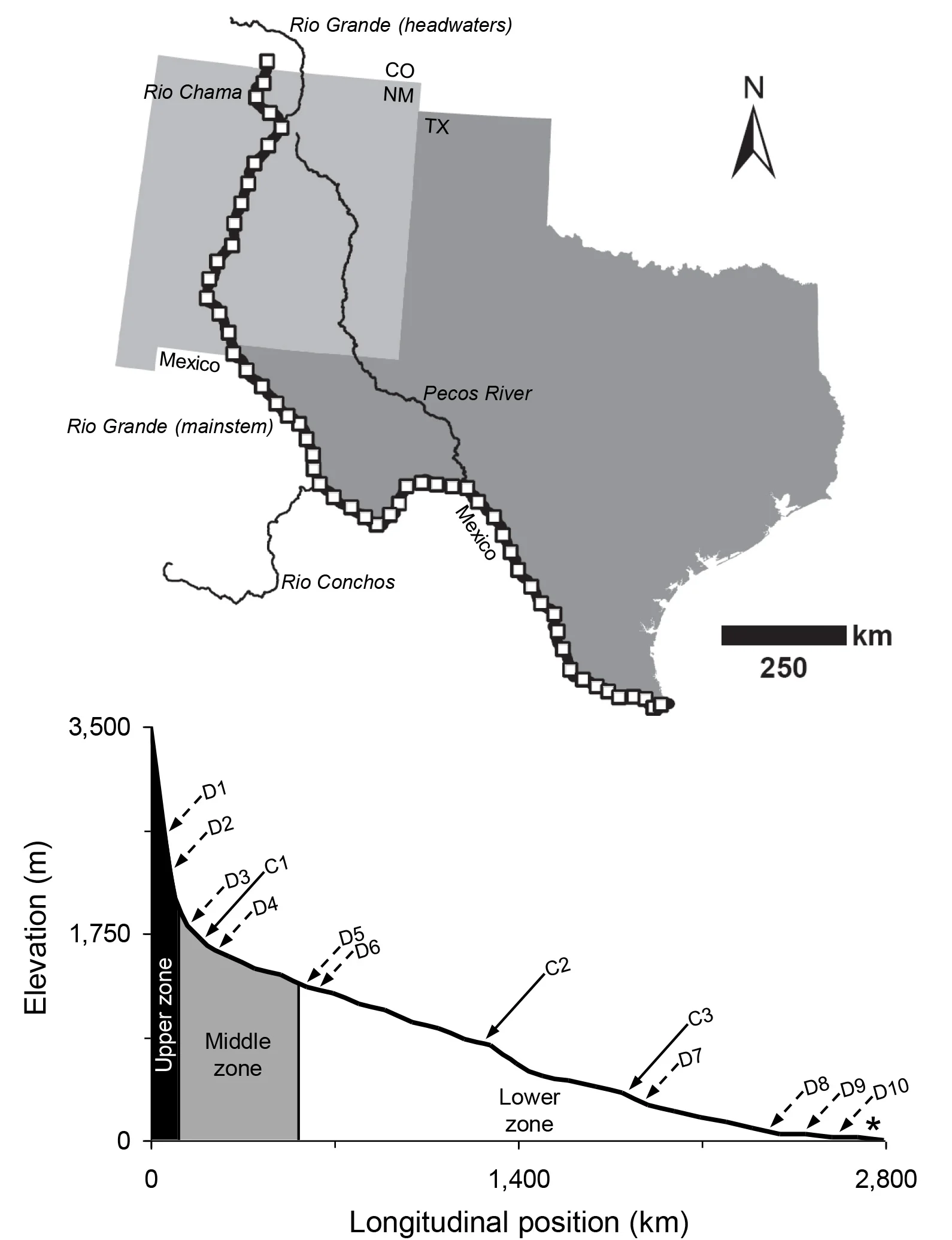
A 2,800 km stretch of the Rio Chama/Rio Grande, broken into 50 km segments (delineated by white boxes), is shown in the upper panel. The lower panel shows a longitudinal profile of the same river with distinct 'upper,' 'middle,' and 'lower' ichthyofaunal zones. The longitudinal positions of dams are shown along the river profile by 'D' labels and major river confluences are indicated by 'C' labels. A complete explanation of the figure is provided in McGarvey (2011).
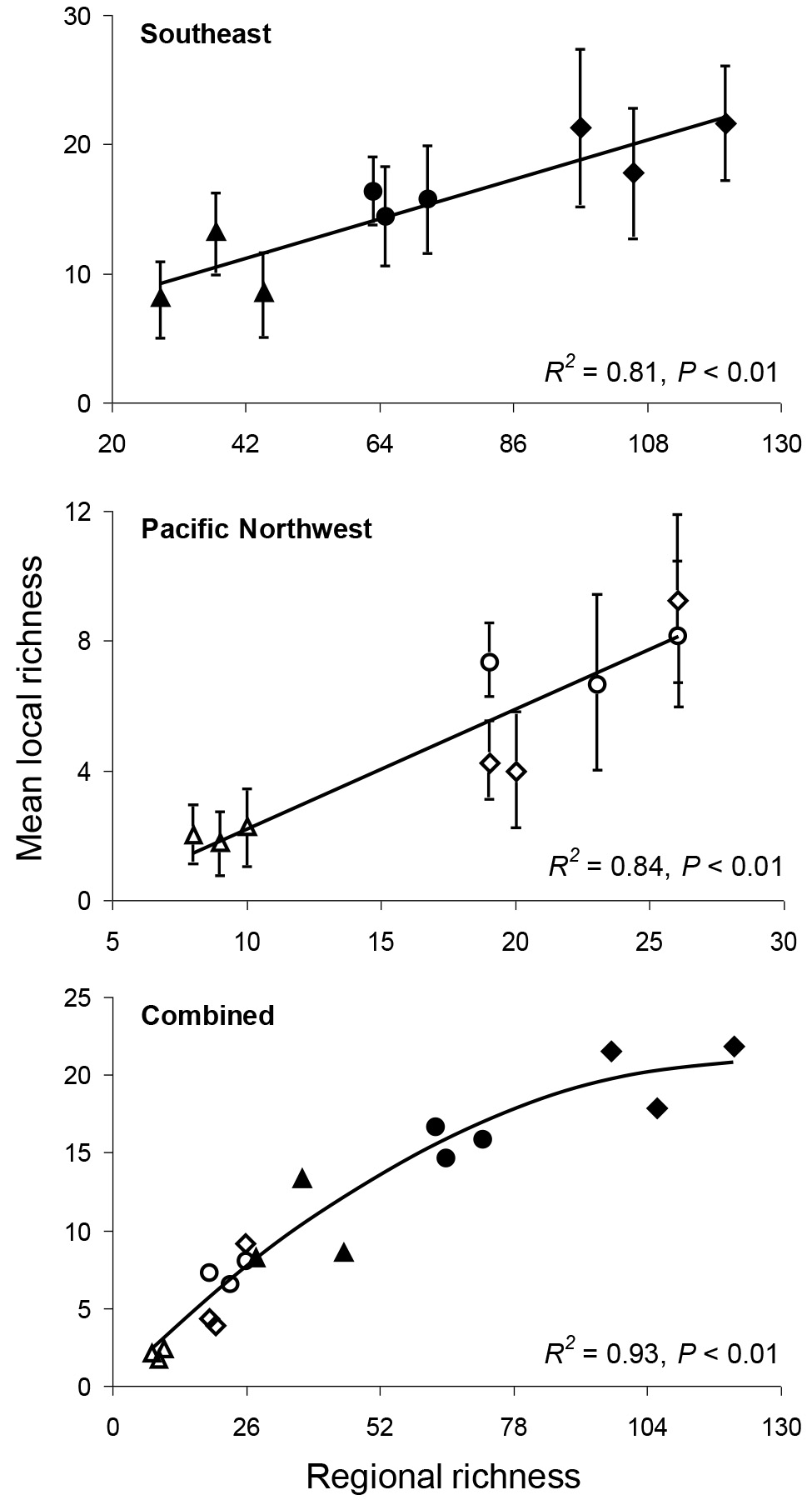
Having articulated an operational, biologically meaningful classification of RSP's for river fishes, I set about using them in tests of freshwater macroecology and community structure. One example of the kind of novel insight that may be possible by using ichthyofaunal zones as RSP's is a zonal test of 'community saturation.' Ecological theory posits that as communities become more diverse, they may approach a saturation point at which it is difficult or impossible for a new species to become locally established without driving a resident species out ('biotic resistance'). An early and widely used test of the saturation hypothesis was to plot regional richness (i.e., the number of species comprising the RSP) against the average local richness of a site (i.e., the number of species that tend to co-occur or interact on a real-time, daily basis) within each region. Plots that reached or approached an asymptote were thought to reflect local saturation, while plots that remained linear suggested that local communities may be nothing more than random samples from their respective RSP's. (More recently, the statistical limitations of the 'local vs. regional richness' method have been recognized. But a log-ratio transformation has also been shown to correct for these statistical problems.)
I used ichthyofaunal zones as RSP's (i.e., regional richness estimates) and the averages of standardized point samples within each zone as local richness estimates to test the saturation hypothesis for Southeast and Pacific Northwest fishes. As shown below, the results provided no evidence of saturation for the Pacific Northwest rivers (consistent with the observation that a disproportionately large number of invasive fishes have become established there), but did indicate that the Southeast rivers may be nearly saturated with species.
Local vs. regional richness plots are shown at right for ichthyofaunal zones in Southeast and Pacific Northwest rivers. Local richness values are the averages of multiple, independent samples collected within each region or zone, with vertical errors bars depicting variation in local richness (+/- 1 S.D.). No evidence of curvature (i.e., an asymptote) was detected for the Pacific Northwest data while moderate curvature was detected for the Southeast data (asymptotic line not shown). However, when the two biogeographic regions are combined in a single plot (at bottom), a strongly asymptotic relationship between local and regional richness becomes apparent.